Длиннонос бесхвостый трубкогубый - Anoura fistulata вид
Видеофрагменты

Warning: Invalid argument supplied for foreach() in /var/www/u0014601/data/www/clasbio.ru/classification.php on line 1798
Количество видов в «сестринских» таксонах
Эволюционный успех ян-рукокрылых объяснился открытым каналом Розенталя
Долгое время зоологи разделяли отряд рукокрылых (Chiroptera) на две основные группы: летучих мышей (Microchiroptera) и крыланов (Megachiroptera). Первые отличаются относительно мелкими размерами и активно используют эхолокацию, чтобы ориентироваться в пространстве и охотиться. Вторые крупнее и, предпочитают эхолокации зрение. Такая классификация кажется очень логичной. Однако генетические исследования, проведенные в начале нынешнего века, показали, что она неверна. Согласно современным представлениям, крыланы вместе с пятью семействами летучих мышей, включая подковоносов (Rhinolophidae) и ложных вампиров (Megadermatidae), формируют один подотряд (Yinpterochiroptera), а около пятнадцати остальных семейств летучих мышей — второй (Yangochiroptera). Устоявшихся русских названий для этих групп не существует, но, поскольку их латинские обозначения происходят от заимствованных из древнекитайской философии терминов инь и ян, в теории их можно называть инь-рукокрылыми и ян-рукокрылыми.
Глядя на новое эволюционное древо рукокрылых, трудно однозначно ответить на вопрос, когда эти млекопитающие начали пользоваться эхолокацией. Возможно, данная способность появилась уже у общего предка всех рукокрылых, но крыланы ее утратили. Согласно альтернативной гипотезе, древнейшие рукокрылые не пользовались эхолокацией, а представители подотрядов Yinpterochiroptera (за исключением подавляющего большинства крыланов) и Yangochiroptera приобрели этот навык независимо друг от друга.
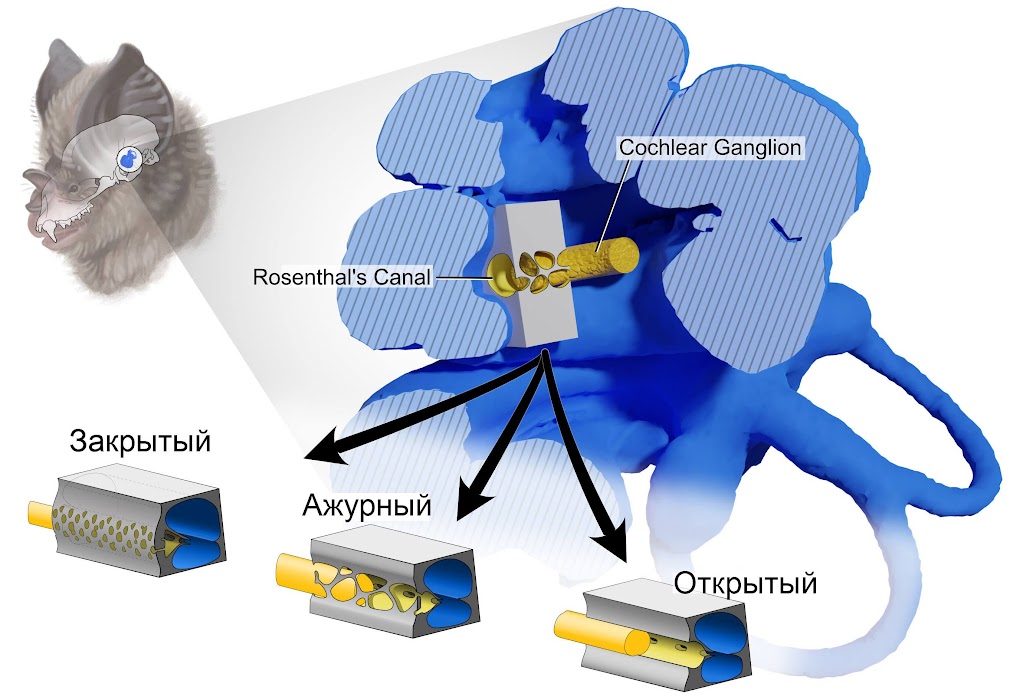
Команда зоологов под руководством Бенджамина Сульсера (R. Benjamin Sulser) из Американского музея естественной истории решила больше узнать об эволюции эхолокации у рукокрылых. Для этого исследователи с помощью компьютерной томографии и гистологического анализа изучили строение внутреннего уха 39 видов летучих мышей и крыланов из 19 семейств. В центре их внимания оказалась структура под названием спиральный ганглий — совокупность тел биполярных нейронов, расположенных в костном стержне улитки. Аксоны этих нейронов формируют кохлеарный нерв, по которому слуховая информация передается от волосковых клеток кортиева органа к кохлеарным ядрам головного мозга.
Спиральный ганглий заключен в костный канал — так называемый канал Розенталя. У подавляющего большинства сумчатых и плацентарных млекопитающих, включая крыланов и летучих мышей из подотряда Yinpterochiroptera, стенки этой структуры толстые, с рядом мелких отверстий на одной из сторон. Через эти отверстия наружу выходят аксоны нейронов, сливающиеся в ствол кохлеарного нерва. Однако у представителей подотряда Yangochiroptera Сульсер и его соавторы обнаружили совершенно иную картину. У некоторых видов этих рукокрылых одна из стенок канала Розенталя частично или почти полностью исчезла, а у других стала ажурной, с многочисленными крупными отверстиями.
Исследователи отмечают, что строение канала Розенталя у Yangochiroptera значительно варьирует от вида к виду. Судя по всему, общий предок данной группы обладал открытым каналом, а затем у отдельных видов из разных семейств возникла ажурная стенка. Представители рода зайцегубов (Noctilio) и вовсе вторично обзавелись закрытым каналом стандартного строения. Напротив, у Yinpterochiroptera строение канала Розенталя очень консервативное и примерно совпадает у разных видов.
От формы канала Розенталя зависят диаметр и плотность спирального ганглия — а значит, и его способность передавать слуховые сигналы. В канале с открытой или ажурной стенкой можно уместить больше нейронов, заложив основу для более качественной эхолокации. Сульсер и его коллеги допускают, что необычное строение канала Розенталя позволило представителям Yangochiroptera освоить различные эхолокационные стратегии и занять многочисленные экологические ниши. Возможно, именно поэтому данная группа достигла большего успеха, чем Yinpterochiroptera: сегодня к ней принадлежит 82 процента всех видов рукокрылых, использующих эхолокацию. Представителям подотряда Yinpterochiroptera, у которых строение канала Розенталая более консервативное, не удалось достичь такого же уровня разнообразия.
К сожалению, полученные результаты не приблизили исследователей к ответу на вопрос, на каком этапе своей истории рукокрылые освоили эхолокацию. Тем не менее, авторы надеются, что, изучая анатомию внутреннего уха ископаемых летучих мышей и крыланов, можно будет пролить свет на эволюцию этой группы.
Сергей Коленов

Длинный язык летучей мыши
На фото трубкогубая нектарная летучая мышь (Anoura fistulata) пьет сладкий сироп из соломинки, любезно предоставленной учеными. Так была измерена длина языка этой летучей мыши — почти 8,5 см, что в 1,5 раза превышает длину ее тела! Это рекорд среди млекопитающих и второе место после хамелеонов среди позвоночных.
A. fistulata была открыта в 2005 году в эквадорских Андах, где живут также два близких вида, с которыми ее раньше путали, — Anoura caudifer и бесхвостый длиннонос (Anoura geoffroyi). A. fistulata обитает в горных лесах Анд и влажных лесах предгорий Западного Эквадора, Центральной Колумбии и Перу, а также на пространствах с аридным и семиаридным климатом на юге Колумбии на высоте от 1175 до 2510 м.
У летучих мышей рода Anoura (греческое ἀν означает «отсутствие», οὐρά — «хвост») хвосты либо полностью отсутствуют, либо они очень короткие (3–7 мм), у A. fistulata хвост длиной 4,9 мм и немного выступает за край межбедренной перепонки. Язык у сородичей в два раза короче, чем у A. fistulata: 3,7 см у A. caudifer и 3,9 см у бесхвостого листоноса. Кроме длинного языка A. fistulata отличается удлиненной нижней губой, которая выступает за верхнюю губу на 3,3–4,8 мм и свернута в трубочку. Эту особенность отражает видовое название: fistula переводится с латыни как «трубка». Длинная губа может быть приспособлением для более эффективного извлечения нектара либо просто служить для размещения удлиненного языка.
У летучих мышей подсемейства Glossophaginae (семейство листоносые), в которое входят представители рода Anoura, максимальная длина языка тесно связана с длиной нёба и нижней челюсти. У нектароядных летучих мышей, как и у других млекопитающих, основание языка совпадает с основанием ротовой полости, язык крепится к подъязычной кости и нижней челюсти. У A. fistulata же язык проходит через шею в грудную область, позади трахеи, так что основание языка находится между сердцем и грудиной. Эта часть языка окружена тканевой муфтой, или язычной трубкой (glossal tube). Такой язычной трубки нет ни у каких других летучих мышей, зато есть у панголина (см. картинку дня Язык панголина). Именно она позволила панголину увеличить длину выдвижной части языка.
Летучие мыши подсемейства Glossophaginae пьют нектар и опыляют сотни видов неотропических растений. Цветы этих растений не приспособлены к каким-то конкретным группам летучих мышей (в отличие от растений, цветки которых адаптированы, например, к длине и кривизне клюва колибри). Возможно, это связано с меньшей вариабельностью морфологии ротовых органов летучих мышей по сравнению с клювами птиц или хоботками насекомых.
Однако максимально длинный язык A. fistulata указывает на существование цветка со столь же длинным венчиком. И такой имеется — это Centropogon nigricans из семейства колокольчиковых с длиной венчика 8–9 см. Его пыльцу находили на мордочках A. fistulata, а видеонаблюдения показали, что никакие другие животные его не посещают, так что A. fistulata — вероятнее всего, единственный опылитель этого растения. Специализация на одном виде опылителей крайне редко встречается у цветковых растений, а C. nigricans — единственный подобный пример среди растений, опыляемых летучими мышами. Сами же A. fistulata питаются нектаром и других растений (из семейств бромелиевых, маркгравиевых, пасленовых), а также дополняют свой рацион насекомыми.
Считается, что именно появление язычной трубки, позволившее летучей мыши увеличить длину выдвижной части языка, послужило предпосылкой для коэволюции языка A. fistulata и цветка C. nigricans. Так и сформировались два удивительных исключения.
Юлия Михневич